
The heart is a myogenic muscular organ found in all animals with a circulatory system (including all vertebrates), that is responsible for pumping blood throughout the blood vessels by repeated, rhythmic contractions. The term cardiac (as in cardiology) means "related to the heart" and comes from the Greek καρδιά, kardia, for "heart".
The vertebrate heart is composed of cardiac muscle, which is an involuntary striated muscle tissue found only in this organ, and connective tissue. The average human heart, beating at 72 beats per minute, will beat approximately 2.5 billion times during an average 66 year lifespan, and weighs approximately 250 to 300 grams (9 to 11 oz) in females and 300 to 350 grams (11 to 12 oz) in males.[1]
In invertebrates that possess a circulatory system, the heart is typically a tube or small sac and pumps fluid that contains water and nutrients such as proteins, fats, and sugars. In insects, the "heart" is often called the dorsal tube and insect "blood" is almost always not oxygenated since they usually respirate (breathe) directly from their body surfaces (internal and external) to air. However, the hearts of some other arthropods (including spiders and crustaceans such as crabs and shrimp) and some other animals pump hemolymph, which contains the copper-based protein hemocyanin as an oxygen transporter similar to the iron-based hemoglobin in red blood cells found in vertebrates.
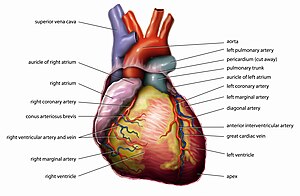
 The human heart
The human heart
From splanchnopleuric mesoderm tissue, the cardiogenic plate develops cranially and laterally to the neural plate. In the cardiogenic plate, two separate angiogenic cell clusters form on either side of the embryo. Each cell cluster coalesces to form an endocardial tube continuous with a dorsal aorta and a vitteloumbilical vein. As embryonic tissue continues to fold, the two endocardial tubes are pushed into the thoracic cavity, begin to fuse together, and complete the fusing process at approximately 21 days.[3]
 At 21 days after conception, the human heart begins beating at 70 to 80 beats per minute and accelerates linearly for the first month of beating.The human embryonic heart begins beating at around 21 days after conception, or five weeks after the last normal menstrual period (LMP). The first day of the LMP is normally used to date the start of the gestation (pregnancy). The human heart begins beating at a rate near the mother’s, about 75-80 beats per minute (BPM).
At 21 days after conception, the human heart begins beating at 70 to 80 beats per minute and accelerates linearly for the first month of beating.The human embryonic heart begins beating at around 21 days after conception, or five weeks after the last normal menstrual period (LMP). The first day of the LMP is normally used to date the start of the gestation (pregnancy). The human heart begins beating at a rate near the mother’s, about 75-80 beats per minute (BPM).
The embryonic heart rate (EHR) then accelerates by approximately 100 BPM during the first month to peak at 165-185 BPM during the early 7th week, (early 9th week after the LMP). This acceleration is approximately 3.3 BPM per day, or about 10 BPM every three days, which is an increase of 100 BPM in the first month.[4] After 9.1 weeks after the LMP, it decelerates to about 152 BPM (+/-25 BPM) during the 15th week post LMP. After the 15th week, the deceleration slows to an average rate of about 145 (+/-25 BPM) BPM, at term. The regression formula, which describes this acceleration before the embryo reaches 25 mm in crown-rump length, or 9.2 LMP weeks, is: the Age in days = EHR(0.3)+6. There is no difference in female and male heart rates before birth.[5]
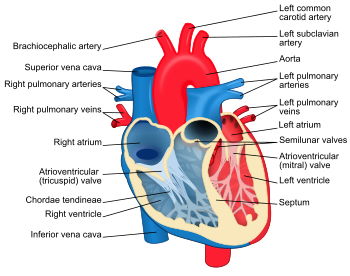 Structure diagram of the human heart from an anterior view. Blue components indicate de-oxygenated blood pathways and red components indicate oxygenated pathways.The human heart has a mass of between 250 and 350 grams and is about the size of a fist.[7] It is located anterior to the vertebral column and posterior to the sternum.
Structure diagram of the human heart from an anterior view. Blue components indicate de-oxygenated blood pathways and red components indicate oxygenated pathways.The human heart has a mass of between 250 and 350 grams and is about the size of a fist.[7] It is located anterior to the vertebral column and posterior to the sternum.
It is enclosed in a double-walled sac called the pericardium. The superficial part of this sac is called the fibrous pericardium. This sac protects the heart, anchors its surrounding structures, and prevents overfilling of the heart with blood.
The outer wall of the human heart is composed of three layers. The outer layer is called the epicardium, or visceral pericardium since it is also the inner wall of the pericardium. The middle layer is called the myocardium and is composed of muscle which contracts. The inner layer is called the endocardium and is in contact with the blood that the heart pumps. Also, it merges with the inner lining (endothelium) of blood vessels and covers heart valves.[8]
The human heart has four chambers, two superior atria and two inferior ventricles. The atria are the receiving chambers and the ventricles are the discharging chambers. The right ventricle discharges into the lungs to oxygenate the blood. The left ventricle discharges its blood toward the rest of the body via the aorta.
The pathway of blood through the human heart consists of a pulmonary circuit and a systemic circuit. Blood flows through the heart in one direction, from the atria to the ventricles, and out of the great arteries, or the aorta for example. This is done by four valves which are the tricuspid valve, the mitral valve, the aortic valve, and the pulmonary valve.[9]
 Schematic of simplified fish heartPrimitive fish have a four-chambered heart, but the chambers are arranged sequentially so that this primitive heart is quite unlike the four-chambered hearts of mammals and birds. The first chamber is the sinus venosus, which collects de-oxygenated blood, from the body, through the hepatic and cardinal veins. From here, blood flows into the atrium and then to the powerful muscular ventricle where the main pumping action will take place. The fourth and final chamber is the conus arteriosus which contains several valves and sends blood to the ventral aorta. The ventral aorta delivers blood to the gills where it is oxygenated and flows, through the dorsal aorta, into the rest of the body. (In tetrapods, the ventral aorta has divided in two; one half forms the ascending aorta, while the other forms the pulmonary artery).[6]
Schematic of simplified fish heartPrimitive fish have a four-chambered heart, but the chambers are arranged sequentially so that this primitive heart is quite unlike the four-chambered hearts of mammals and birds. The first chamber is the sinus venosus, which collects de-oxygenated blood, from the body, through the hepatic and cardinal veins. From here, blood flows into the atrium and then to the powerful muscular ventricle where the main pumping action will take place. The fourth and final chamber is the conus arteriosus which contains several valves and sends blood to the ventral aorta. The ventral aorta delivers blood to the gills where it is oxygenated and flows, through the dorsal aorta, into the rest of the body. (In tetrapods, the ventral aorta has divided in two; one half forms the ascending aorta, while the other forms the pulmonary artery).[6]
In the adult fish, the four chambers are not arranged in a straight row but, instead, form an S-shape with the latter two chambers lying above the former two. This relatively simpler pattern is found in cartilaginous fish and in the ray-finned fish. In teleosts, the conus arteriosus is very small and can more accurately be described as part of the aorta rather than of the heart proper. The conus arteriosus is not present in any amniotes, presumably having been absorbed into the ventricles over the course of evolution. Similarly, while the sinus venosus is present as a vestigial structure in some reptiles and birds, it is otherwise absorbed into the right atrium and is no longer distinguishable.[6]
In living amphibians, the atrium is divided into two separate chambers by the presence of a muscular septum even though there is only a single ventricle. The sinus venosus, which remains large in amphibians but connects only to the right atrium, receives blood from the vena cavae, with the pulmonary vein by-passing it entirely to enter the left atrium.
In the heart of lungfish, the septum extends part-way into the ventricle. This allows for some degree of separation between the de-oxygenated bloodstream destined for the lungs and the oxygenated stream that is delivered to the rest of the body. The absence of such a division in living amphibian species may be at least partly due to the amount of respiration that occurs through the skin in such species; thus, the blood returned to the heart through the vena cavae is, in fact, already partially oxygenated. As a result, there may be less need for a finer division between the two bloodstreams than in lungfish or other tetrapods. Nonetheless, in at least some species of amphibian, the spongy nature of the ventricle seems to maintain more of a separation between the bloodstreams than appears the case at first glance. Furthermore, the conus arteriosus has lost its original valves and contains a spiral valve, instead, that divides it into two parallel parts, thus helping to keep the two bloodstreams separate.[6]
The heart of most reptiles (except for crocodilians; see below) has a similar structure to that of lungfish but, here, the septum is generally much larger. This divides the ventricle into two halves but, because the septum does not reach the whole length of the heart, there is a considerable gap near the openings to the pulmonary artery and the aorta. In practice, however, in the majority of reptilian species, there appears to be little, if any, mixing between the bloodstreams, so the aorta receives, essentially, only oxygenated blood.[6]
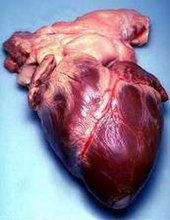 Human heart removed from a 64-year-old male
Human heart removed from a 64-year-old male  Surface anatomy of the human heart. The heart is demarcated by:
Surface anatomy of the human heart. The heart is demarcated by:
-A point 9 cm to the left of the midsternal line (apex of the heart)
-The seventh right sternocostal articulation
-The upper border of the third right costal cartilage 1 cm from the right sternal line
-The lower border of the second left costal cartilage 2.5 cm from the left lateral sternal line.[10]Archosaurs, (crocodilians, birds), and mammals show complete separation of the heart into two pumps for a total of four heart chambers; it is thought that the four-chambered heart of archosaurs evolved independently from that of mammals. In crocodilians, there is a small opening, the foramen of Panizza, at the base of the arterial trunks and there is some degree of mixing between the blood in each side of the heart; thus, only in birds and mammals are the two streams of blood - those to the pulmonary and systemic circulations - kept entirely separate by a physical barrier.[6]
In the human body, the heart is usually situated in the middle of the thorax with the largest part of the heart slightly offset to the left, although sometimes it is on the right (see dextrocardia), underneath the sternum. The heart is usually felt to be on the left side because the left heart (left ventricle) is stronger (it pumps to all body parts). The left lung is smaller than the right lung because the heart occupies more of the left hemithorax. The heart is fed by the coronary circulation and is enclosed by a sac known as the pericardium; it is also surrounded by the lungs. The pericardium comprises two parts: the fibrous pericardium, made of dense fibrous connective tissue, and a double membrane structure (parietal and visceral pericardium) containing a serous fluid to reduce friction during heart contractions. The heart is located in the mediastinum, which is the central sub-division of the thoracic cavity. The mediastinum also contains other structures, such as the esophagus and trachea, and is flanked on either side by the right and left pulmonary cavities; these cavities house the lungs.[11]
The apex is the blunt point situated in an inferior (pointing down and left) direction. A stethoscope can be placed directly over the apex so that the beats can be counted. It is located posterior to the 5th intercostal space just medial of the left mid-clavicular line. In normal adults, the mass of the heart is 250-350 g (9-12 oz), or about twice the size of a clenched fist (it is about the size of a clenched fist in children), but an extremely diseased heart can be up to 1000 g (2 lb) in mass due to hypertrophy. It consists of four chambers, the two upper atria and the two lower ventricles.
 Blood flow diagram of the human heart. Blue components indicate de-oxygenated blood pathways and red components indicate oxygenated pathways.
Blood flow diagram of the human heart. Blue components indicate de-oxygenated blood pathways and red components indicate oxygenated pathways.  Image showing the conduction system of the heartIn mammals, the function of the right side of the heart (see right heart) is to collect de-oxygenated blood, in the right atrium, from the body (via superior and inferior vena cavae) and pump it, through the tricuspid valve, via the right ventricle, into the lungs (pulmonary circulation) so that carbon dioxide can be dropped off and oxygen picked up (gas exchange). This happens through the passive process of diffusion. The left side (see left heart) collects oxygenated blood from the lungs into the left atrium. From the left atrium the blood moves to the left ventricle, through the bicuspid valve, which pumps it out to the body (via the aorta). On both sides, the lower ventricles are thicker and stronger than the upper atria. The muscle wall surrounding the left ventricle is thicker than the wall surrounding the right ventricle due to the higher force needed to pump the blood through the systemic circulation.
Image showing the conduction system of the heartIn mammals, the function of the right side of the heart (see right heart) is to collect de-oxygenated blood, in the right atrium, from the body (via superior and inferior vena cavae) and pump it, through the tricuspid valve, via the right ventricle, into the lungs (pulmonary circulation) so that carbon dioxide can be dropped off and oxygen picked up (gas exchange). This happens through the passive process of diffusion. The left side (see left heart) collects oxygenated blood from the lungs into the left atrium. From the left atrium the blood moves to the left ventricle, through the bicuspid valve, which pumps it out to the body (via the aorta). On both sides, the lower ventricles are thicker and stronger than the upper atria. The muscle wall surrounding the left ventricle is thicker than the wall surrounding the right ventricle due to the higher force needed to pump the blood through the systemic circulation.
Starting in the right atrium, the blood flows through the tricuspid valve to the right ventricle. Here, it is pumped out the pulmonary semilunar valve and travels through the pulmonary artery to the lungs. From there, oxygenated blood flows back through the pulmonary vein to the left atrium. It then travels through the mitral valve to the left ventricle, from where it is pumped through the aortic semilunar valve to the aorta. The aorta forks and the blood is divided between major arteries which supply the upper and lower body. The blood travels in the arteries to the smaller arterioles and then, finally, to the tiny capillaries which feed each cell. The (relatively) deoxygenated blood then travels to the venules, which coalesce into veins, then to the inferior and superior venae cavae and finally back to the right atrium where the process began.
The heart is effectively a syncytium, a meshwork of cardiac muscle cells interconnected by contiguous cytoplasmic bridges. This relates to electrical stimulation of one cell spreading to neighboring cells.
Some cardiac cells are self-excitable, contracting without any signal from the nervous system, even if removed from the heart and placed in culture. Each of these cells have their own intrinsic contraction rhythm. A region of the human heart called the sinoatrial node, or pacemaker, sets the rate and timing at which all cardiac muscle cells contract. The SA node generates electrical impulses, much like those produced by nerve cells. Because cardiac muscle cells are electrically coupled by inter-calated disks between adjacent cells, impulses from the SA node spread rapidly through the walls of the artria, causing both artria to contract in unison. The impulses also pass to another region of specialized cardiac muscle tissue, a relay point called the atrioventricular node, located in the wall between the right atrium and the right ventricle. Here, the impulses are delayed for about 0.1s before spreading to the walls of the ventricle. The delay ensures that the artria empty completely before the ventricles contract. Specialized muscle fibers called Purkinje fibers then conduct the signals to the apex of the heart along and throughout the ventricular walls. The Purkinje fibres form conducting pathways called bundle branches. This entire cycle, a single heart beat, lasts about 0.8 seconds. The impulses generated during the heart cycle produce electrical currents, which are conducted through body fluids to the skin, where they can be detected by electrodes and recorded as an electrocardiogram (ECG or EKG).[12] The events related to the flow or blood pressure that occurs from the beginning of one heartbeat to the beginning of the next can be referred to a cardiac cycle.[13]
The SA node is found in all amniotes but not in more primitive vertebrates. In these animals, the muscles of the heart are relatively continuous and the sinus venosus coordinates the beat which passes in a wave through the remaining chambers. Indeed, since the sinus venosus is incorporated into the right atrium in amniotes, it is likely homologous with the SA node. In teleosts, with their vestigial sinus venosus, the main centre of coordination is, instead, in the atrium. The rate of heartbeat varies enormously between different species, ranging from around 20 beats per minute in codfish to around 600 in hummingbirds.[6]
Cardiac arrest is the sudden cessation of normal heart rhythm which can include a number of pathologies such as tachycardia, an extremely rapid heart beat which prevents the heart from effectively pumping blood, fibrillation, which is an irregular and ineffective heart rhythm, and asystole, which is the cessation of heart rhythm entirely.
Cardiac tamponade is a condition in which the fibrous sac surrounding the heart fills with excess fluid or blood, suppressing the heart's ability to beat properly. Tamponade is treated by pericardiocentesis, the gentle insertion of the needle of a syringe into the pericardial sac (avoiding the heart itself) on an angle, usually from just below the sternum, and gently withdrawing the tamponading fluids.
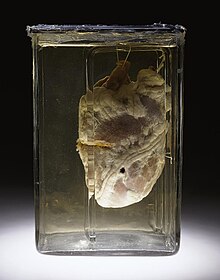 A preserved human heart with a visible gunshot woundThe valves of the heart were discovered by a physician of the Hippocratean school around the 4th century BC. However, their function was not properly understood then. Because blood pools in the veins after death, arteries look empty. Ancient anatomists assumed they were filled with air and that they were for transport of air.
A preserved human heart with a visible gunshot woundThe valves of the heart were discovered by a physician of the Hippocratean school around the 4th century BC. However, their function was not properly understood then. Because blood pools in the veins after death, arteries look empty. Ancient anatomists assumed they were filled with air and that they were for transport of air.
Philosophers distinguished veins from arteries, but thought the pulse was a property of arteries themselves. Erasistratos observed the arteries that were cut during life bleed. He ascribed the fact to the phenomenon that air escaping from an artery is replaced with blood which entered by very small vessels between veins and arteries. Thus he apparently postulated capillaries, but with reversed flow of blood.
The 2nd century AD, Greek physician Galenos (Galen) knew blood vessels carried blood and identified venous (dark red) and arterial (brighter and thinner) blood, each with distinct and separate functions. Growth and energy were derived from venous blood created in the liver from chyle, while arterial blood gave vitality by containing pneuma (air) and originated in the heart. Blood flowed from both creating organs to all parts of the body, where it was consumed and there was no return of blood to the heart or liver. The heart did not pump blood around, the heart's motion sucked blood in during diastole and the blood moved by the pulsation of the arteries themselves.
Galen believed the arterial blood was created by venous blood passing from the left ventricle to the right through 'pores' in the interventricular septum, while air passed from the lungs via the pulmonary artery to the left side of the heart. As the arterial blood was created, 'sooty' vapors were created and passed to the lungs, also via the pulmonary artery, to be exhaled.
The vertebrate heart is composed of cardiac muscle, which is an involuntary striated muscle tissue found only in this organ, and connective tissue. The average human heart, beating at 72 beats per minute, will beat approximately 2.5 billion times during an average 66 year lifespan, and weighs approximately 250 to 300 grams (9 to 11 oz) in females and 300 to 350 grams (11 to 12 oz) in males.[1]
In invertebrates that possess a circulatory system, the heart is typically a tube or small sac and pumps fluid that contains water and nutrients such as proteins, fats, and sugars. In insects, the "heart" is often called the dorsal tube and insect "blood" is almost always not oxygenated since they usually respirate (breathe) directly from their body surfaces (internal and external) to air. However, the hearts of some other arthropods (including spiders and crustaceans such as crabs and shrimp) and some other animals pump hemolymph, which contains the copper-based protein hemocyanin as an oxygen transporter similar to the iron-based hemoglobin in red blood cells found in vertebrates.

 | |
| Problems listening to this file? See media help. | |
Contents |
Early development
Main article: Heart development
The mammalian heart is derived from embryonic mesoderm germ-layer cells that differentiate after gastrulation into mesothelium, endothelium, and myocardium. Mesothelial pericardium forms the outer lining of the heart. The inner lining of the heart, lymphatic and blood vessels, develop from endothelium. Heart muscle is termed myocardium.[2]From splanchnopleuric mesoderm tissue, the cardiogenic plate develops cranially and laterally to the neural plate. In the cardiogenic plate, two separate angiogenic cell clusters form on either side of the embryo. Each cell cluster coalesces to form an endocardial tube continuous with a dorsal aorta and a vitteloumbilical vein. As embryonic tissue continues to fold, the two endocardial tubes are pushed into the thoracic cavity, begin to fuse together, and complete the fusing process at approximately 21 days.[3]

The embryonic heart rate (EHR) then accelerates by approximately 100 BPM during the first month to peak at 165-185 BPM during the early 7th week, (early 9th week after the LMP). This acceleration is approximately 3.3 BPM per day, or about 10 BPM every three days, which is an increase of 100 BPM in the first month.[4] After 9.1 weeks after the LMP, it decelerates to about 152 BPM (+/-25 BPM) during the 15th week post LMP. After the 15th week, the deceleration slows to an average rate of about 145 (+/-25 BPM) BPM, at term. The regression formula, which describes this acceleration before the embryo reaches 25 mm in crown-rump length, or 9.2 LMP weeks, is: the Age in days = EHR(0.3)+6. There is no difference in female and male heart rates before birth.[5]
Structure
The structure of the heart varies among the different branches of the animal kingdom. (See Circulatory system.) Cephalopods have two "gill hearts" and one "systemic heart". In vertebrates, the heart lies in the anterior part of the body cavity, dorsal to the gut. It is always surrounded by a pericardium, which is usually a distinct structure, but may be continuous with the peritoneum in jawless and cartilaginous fish. Hagfishes, uniquely among vertebrates, also possess a second heart-like structure in the tail.[6]In humans
Main article: Human heart

It is enclosed in a double-walled sac called the pericardium. The superficial part of this sac is called the fibrous pericardium. This sac protects the heart, anchors its surrounding structures, and prevents overfilling of the heart with blood.
The outer wall of the human heart is composed of three layers. The outer layer is called the epicardium, or visceral pericardium since it is also the inner wall of the pericardium. The middle layer is called the myocardium and is composed of muscle which contracts. The inner layer is called the endocardium and is in contact with the blood that the heart pumps. Also, it merges with the inner lining (endothelium) of blood vessels and covers heart valves.[8]
The human heart has four chambers, two superior atria and two inferior ventricles. The atria are the receiving chambers and the ventricles are the discharging chambers. The right ventricle discharges into the lungs to oxygenate the blood. The left ventricle discharges its blood toward the rest of the body via the aorta.
The pathway of blood through the human heart consists of a pulmonary circuit and a systemic circuit. Blood flows through the heart in one direction, from the atria to the ventricles, and out of the great arteries, or the aorta for example. This is done by four valves which are the tricuspid valve, the mitral valve, the aortic valve, and the pulmonary valve.[9]
In fish

In the adult fish, the four chambers are not arranged in a straight row but, instead, form an S-shape with the latter two chambers lying above the former two. This relatively simpler pattern is found in cartilaginous fish and in the ray-finned fish. In teleosts, the conus arteriosus is very small and can more accurately be described as part of the aorta rather than of the heart proper. The conus arteriosus is not present in any amniotes, presumably having been absorbed into the ventricles over the course of evolution. Similarly, while the sinus venosus is present as a vestigial structure in some reptiles and birds, it is otherwise absorbed into the right atrium and is no longer distinguishable.[6]
In double circulatory systems
In amphibians and most reptiles, a double circulatory system is used but the heart is not completely separated into two pumps. The development of the double system is necessitated by the presence of lungs which deliver oxygenated blood directly to the heart.In living amphibians, the atrium is divided into two separate chambers by the presence of a muscular septum even though there is only a single ventricle. The sinus venosus, which remains large in amphibians but connects only to the right atrium, receives blood from the vena cavae, with the pulmonary vein by-passing it entirely to enter the left atrium.
In the heart of lungfish, the septum extends part-way into the ventricle. This allows for some degree of separation between the de-oxygenated bloodstream destined for the lungs and the oxygenated stream that is delivered to the rest of the body. The absence of such a division in living amphibian species may be at least partly due to the amount of respiration that occurs through the skin in such species; thus, the blood returned to the heart through the vena cavae is, in fact, already partially oxygenated. As a result, there may be less need for a finer division between the two bloodstreams than in lungfish or other tetrapods. Nonetheless, in at least some species of amphibian, the spongy nature of the ventricle seems to maintain more of a separation between the bloodstreams than appears the case at first glance. Furthermore, the conus arteriosus has lost its original valves and contains a spiral valve, instead, that divides it into two parallel parts, thus helping to keep the two bloodstreams separate.[6]
The heart of most reptiles (except for crocodilians; see below) has a similar structure to that of lungfish but, here, the septum is generally much larger. This divides the ventricle into two halves but, because the septum does not reach the whole length of the heart, there is a considerable gap near the openings to the pulmonary artery and the aorta. In practice, however, in the majority of reptilian species, there appears to be little, if any, mixing between the bloodstreams, so the aorta receives, essentially, only oxygenated blood.[6]
The fully divided heart


-A point 9 cm to the left of the midsternal line (apex of the heart)
-The seventh right sternocostal articulation
-The upper border of the third right costal cartilage 1 cm from the right sternal line
-The lower border of the second left costal cartilage 2.5 cm from the left lateral sternal line.[10]
In the human body, the heart is usually situated in the middle of the thorax with the largest part of the heart slightly offset to the left, although sometimes it is on the right (see dextrocardia), underneath the sternum. The heart is usually felt to be on the left side because the left heart (left ventricle) is stronger (it pumps to all body parts). The left lung is smaller than the right lung because the heart occupies more of the left hemithorax. The heart is fed by the coronary circulation and is enclosed by a sac known as the pericardium; it is also surrounded by the lungs. The pericardium comprises two parts: the fibrous pericardium, made of dense fibrous connective tissue, and a double membrane structure (parietal and visceral pericardium) containing a serous fluid to reduce friction during heart contractions. The heart is located in the mediastinum, which is the central sub-division of the thoracic cavity. The mediastinum also contains other structures, such as the esophagus and trachea, and is flanked on either side by the right and left pulmonary cavities; these cavities house the lungs.[11]
The apex is the blunt point situated in an inferior (pointing down and left) direction. A stethoscope can be placed directly over the apex so that the beats can be counted. It is located posterior to the 5th intercostal space just medial of the left mid-clavicular line. In normal adults, the mass of the heart is 250-350 g (9-12 oz), or about twice the size of a clenched fist (it is about the size of a clenched fist in children), but an extremely diseased heart can be up to 1000 g (2 lb) in mass due to hypertrophy. It consists of four chambers, the two upper atria and the two lower ventricles.
Functioning


Starting in the right atrium, the blood flows through the tricuspid valve to the right ventricle. Here, it is pumped out the pulmonary semilunar valve and travels through the pulmonary artery to the lungs. From there, oxygenated blood flows back through the pulmonary vein to the left atrium. It then travels through the mitral valve to the left ventricle, from where it is pumped through the aortic semilunar valve to the aorta. The aorta forks and the blood is divided between major arteries which supply the upper and lower body. The blood travels in the arteries to the smaller arterioles and then, finally, to the tiny capillaries which feed each cell. The (relatively) deoxygenated blood then travels to the venules, which coalesce into veins, then to the inferior and superior venae cavae and finally back to the right atrium where the process began.
The heart is effectively a syncytium, a meshwork of cardiac muscle cells interconnected by contiguous cytoplasmic bridges. This relates to electrical stimulation of one cell spreading to neighboring cells.
Some cardiac cells are self-excitable, contracting without any signal from the nervous system, even if removed from the heart and placed in culture. Each of these cells have their own intrinsic contraction rhythm. A region of the human heart called the sinoatrial node, or pacemaker, sets the rate and timing at which all cardiac muscle cells contract. The SA node generates electrical impulses, much like those produced by nerve cells. Because cardiac muscle cells are electrically coupled by inter-calated disks between adjacent cells, impulses from the SA node spread rapidly through the walls of the artria, causing both artria to contract in unison. The impulses also pass to another region of specialized cardiac muscle tissue, a relay point called the atrioventricular node, located in the wall between the right atrium and the right ventricle. Here, the impulses are delayed for about 0.1s before spreading to the walls of the ventricle. The delay ensures that the artria empty completely before the ventricles contract. Specialized muscle fibers called Purkinje fibers then conduct the signals to the apex of the heart along and throughout the ventricular walls. The Purkinje fibres form conducting pathways called bundle branches. This entire cycle, a single heart beat, lasts about 0.8 seconds. The impulses generated during the heart cycle produce electrical currents, which are conducted through body fluids to the skin, where they can be detected by electrodes and recorded as an electrocardiogram (ECG or EKG).[12] The events related to the flow or blood pressure that occurs from the beginning of one heartbeat to the beginning of the next can be referred to a cardiac cycle.[13]
The SA node is found in all amniotes but not in more primitive vertebrates. In these animals, the muscles of the heart are relatively continuous and the sinus venosus coordinates the beat which passes in a wave through the remaining chambers. Indeed, since the sinus venosus is incorporated into the right atrium in amniotes, it is likely homologous with the SA node. In teleosts, with their vestigial sinus venosus, the main centre of coordination is, instead, in the atrium. The rate of heartbeat varies enormously between different species, ranging from around 20 beats per minute in codfish to around 600 in hummingbirds.[6]
Cardiac arrest is the sudden cessation of normal heart rhythm which can include a number of pathologies such as tachycardia, an extremely rapid heart beat which prevents the heart from effectively pumping blood, fibrillation, which is an irregular and ineffective heart rhythm, and asystole, which is the cessation of heart rhythm entirely.
Cardiac tamponade is a condition in which the fibrous sac surrounding the heart fills with excess fluid or blood, suppressing the heart's ability to beat properly. Tamponade is treated by pericardiocentesis, the gentle insertion of the needle of a syringe into the pericardial sac (avoiding the heart itself) on an angle, usually from just below the sternum, and gently withdrawing the tamponading fluids.
History of discoveries

Philosophers distinguished veins from arteries, but thought the pulse was a property of arteries themselves. Erasistratos observed the arteries that were cut during life bleed. He ascribed the fact to the phenomenon that air escaping from an artery is replaced with blood which entered by very small vessels between veins and arteries. Thus he apparently postulated capillaries, but with reversed flow of blood.
The 2nd century AD, Greek physician Galenos (Galen) knew blood vessels carried blood and identified venous (dark red) and arterial (brighter and thinner) blood, each with distinct and separate functions. Growth and energy were derived from venous blood created in the liver from chyle, while arterial blood gave vitality by containing pneuma (air) and originated in the heart. Blood flowed from both creating organs to all parts of the body, where it was consumed and there was no return of blood to the heart or liver. The heart did not pump blood around, the heart's motion sucked blood in during diastole and the blood moved by the pulsation of the arteries themselves.
Galen believed the arterial blood was created by venous blood passing from the left ventricle to the right through 'pores' in the interventricular septum, while air passed from the lungs via the pulmonary artery to the left side of the heart. As the arterial blood was created, 'sooty' vapors were created and passed to the lungs, also via the pulmonary artery, to be exhaled.
No comments:
Post a Comment